
In alcuni vecchi articoli avevo già accennato l’argomento ma senza mai dilungarmi a sufficienza, oggi invece entrerò un po’ più nel dettaglio.

Il nostro corpo è composto da tessuti, per la precisione da cinque diversi tipi di tessuti, uno di questi è quello muscolare.
I tessuti muscolari sono costituiti da cellule di forma allungata, organizzate in strutture chiamate miofibrille, la cui caratteristica principale è quella della contrattilità.
I muscoli si dividono in muscoli volontari e muscoli involontari (o vegetativi) non dipendenti dalla volontà dell’uomo ma primari ai fini della sopravvivenza (es. cuore). Neanche i tessuti muscolari sono tutti uguali, essi, infatti, si dividono in lisci e striati. Nei primi, la forma delle cellule è affusolata, interdigitata tra di loro fino a formare lamine di muscolatura nelle pareti di organi cavi. Per questa caratteristica i muscoli lisci hanno la possibilità di effettuare contrazioni ritmiche, alternate a fasi rilassamento. Un’altra caratteristica dei muscoli lisci è, oltre a quella di ipertrofizzarsi, quella di moltiplicarsi (es. fibrocellule dell’utero durante la gravidanza).
I muscoli scheletrici volontari fanno parte dell’apparato locomotore e permettono il movimento.
La forza di un muscolo dipende da diversi fattori: la superficie della sua sezione trasversale, la sua capacità di attivazione, qualità delle fibre muscolari, coordinazione intramuscolare, coordinaione intermuscolare, vari adattamenti nervosi e altro microfattori.
I dati che oggigiorno si hanno, sembrano confermare che questi tre fattori vengano influenzati, in diversa misura, dall’allenamento.
Ogni muscolo scheletrico è costituito da fasci di fibre, separati da una guaina connettivale e disposti parallelamente gli uni agli altri. Ogni fibra contenuta in questi fasci non è altro che una cellula molto allungata, con diverse centinaia di nuclei.

Epimisio: tessuto connettivo di protezione esterna
Permisio: avvolge i fascicoli (fasci di fibre)
Fibre muscolari: singole cellule muscolari
Endomisio: guaina di tessuto connettivo che avvolge le fibre muscolari.
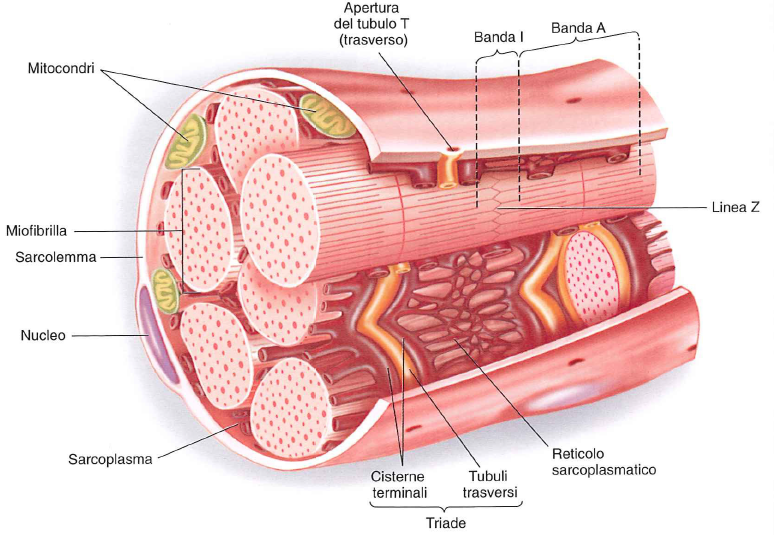
Le fibre muscolari sono praticamente invisibili ad occhio nudo (diametro 10-80 µm). La singola fibra muscolare è avvolta da una membrana plasmatica, il sarcolemma, che racchiude il citoplasma (sarcoplasma). Questa sostanza, contiene filamenti ancor più piccoli: le miofibrille, ovvero elementi contrattili del muscolo scheletrico; lunghi filamenti composti a loro volta da elementi proteici allineati chiamati sarcomeri.
Il sarcoplasma continene mitocondri, ribosomi, glicogeno, fosfocreatina (PC), l’ATP e i suoi precursori (AMP e ADP), lipidi e inoltre accoglie una vasta rete di tubuli trasversi (tubuli T) che sono estensioni del sarcolemma e attraversano la fibra muscolare lateralmente.
All’interno del reticolo endoplasmatico si trova anche una rete longitudinale di tubuli denominata reticolo sarcoplasmatico (RS).
Questi canali membranosi si estendono parallelamente alle miofibrille che circondano completamente. Esso serve a immagazzinare il calcio, micronutriente fondamentale per le contrazioni muscolari.
I sarcomeri, in fila, delle miofibrille sono unità alle estremità dalla “linea Z”.
Un sarcomero contiene gli elementi compresi fra due linee Z nel seguente ordine:
- una banda I (zona chiara)
- una banda A (zona scura)
- una zona H (al centro della banda A)
- la restante parte della banda A
- una seconda banda I
All’interno di una miofibrilla, disposti fianco a fianco, possiamo distinguere due tipi di piccoli filamenti proteici, responsabili dell’azione muscolare, quelli di actina (più sottili, 3000 circa) e miosina (più spessi, 1500). La loro alternanza determina le classiche striature visibili nelle illustrazioni dei muscoli (o al microscopio).
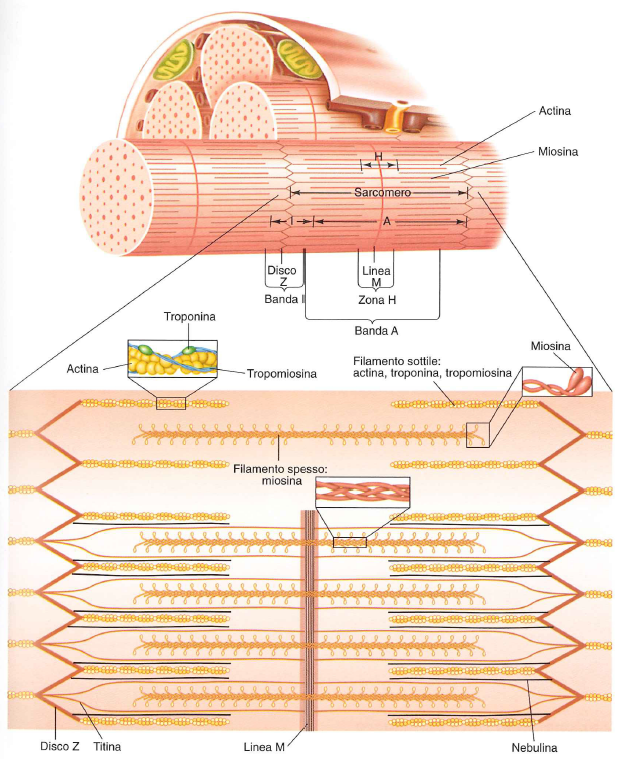
- Banda chiara I: filamenti di actina
- Banda scura A: filamenti di actina e miosina
- Banda H: filamenti di miosina
La banda H si nota solo quando il sarcomero è a riposo perchè i sarcomeri si accorciano durante la contrazione e i filamenti di actina vengono tirati verso l’interno di questa zona che quindi non appare diversa dalla parte restante della banda A, se non per il colore un po’ più chiaro a causa dell’assenza di filamenti di actina.
Nel sarcoplasma, inoltre, sono contenuti i mitocondri: questi potrebbero essere paragonati a minuscole caldaie in cui avvengono le reazioni chimiche di produzione di energia che richiedono ossigeno (dette quindi aerobiche).
Nei mitocondri si ossidano gli acidi grassi e il loro numero è modificabile fino alla fine dell’adolescenza e inizio della giovinezza (17-21), dopo è solo modificabile, entro un certo limite, la loro dimensione. L’ipertrofia mitocondriale si può ottenere grazie ad un’alimentazione sana e ad una buona attività aerobica.
In sintesi, all’interno di ogni fibra muscolare, abbiamo un gran numero di unità contrattili (le miofibrille innervate dai motoneuroni collegati al sistema nevoso centrale) e un gran numero di “centrali energetiche” (i mitocondri), in grado di convertire l’energia chimica derivante dalla trasformazione finale degli alimenti, in energia meccanica (contrazioni muscolari) e quindi in movimento.
Per fare delle proporzioni, quelle che seguono sono le componenti principali del muscolo (espresse in percentuale).
Miofibrille: 20-30%
Sarcoplasma: 20-30%
Mitocondri:15-25%
Depositi di grasso: 10-15%
Altre sostanze sub-cellulari: 4-7%
Capillari: 3-5%
Glicogeno: 2-5%
Tessuto connettivo: 2-3%
La contrazione dei muscoli scheletrici deputati al movimento è innescata dalla volontà di eseguire un gesto: questo impulso parte dal sistema nervoso centrale (SNC) e raggiunge le fibre muscolari tramite le fibre nervose provenienti dai rispettivi motoneuroni, ciascuno dei quali innerva più fibre.
Durante la fase di contrazione, il piccolo accorciamento simultaneo di ciascun sarcomero determina un più sensibile accorciamento della fibra in toto e conseguentemente di tutto il muscolo.
Il controllo della motricità è possibile perchè le numerosissimi fibre che compongono i muscoli vengono attivate in modo selettivo durante le varie contrazioni. La loro immensa “popolazione”, infatti, è suddivisa in unità funzionali, le cosiddette unità motorie.
L’unità motoria, protagonista della contrazione muscolare, è data dall’associazione di un motoneurone con un numero variabile di fibre muscolari (le dimensioni dell’um, quindi, possono essere molto diverse).
Poichè le fibre possono avere una diversa soglia di eccitabilità, uno stesso motoneurone può sollecitare solo alcune fibre dell’unità motoria, oppure tutte le fibre, a seconda dell’intensità dello stimolo ricevuto. Questo aspetto è fondamentale, poichè le singole fibre o si contraggono completamente o non si contraggono affatto (“tutto o nulla”); se non ci fossero le um, il muscolo sarebbe o completamente contratto o completamente rilassato.
Alcune gravi patologie (es. paresi spastica), mostrano come la disorganizzazione di questo sistema possa inibire la motricità o renderla incontrollata.
La possibilità di sollecitare in modo diversificato le um permette di graduare il livello di contrazione da un minimo ad un massimo, mediante l’attivazione di un numero crescente di fibre muscolari.
A seconda della qualità del motoneurone e delle fibre che compongono l’um, la forza prodotta avrà caratteristiche diverse: si svilupperà con una certa rapidità, con un livello più o meno elevato e sarà in grado di mantenere questa contrazione per un periodo maggiore o minore.
I motoneuroni fasici producono contrazioni rapide, di breve durata. I motoneuroni tonici, al contrario, stimolano contrazioni lente e prolungate.
Tipi di fibre
In base alla bibliografia internazionale, possiamo classificare le fibre muscolari in:
- Fibre di lente (tipo I, rosse), ovvero slow twitch oxidative fibers”. Hanno una piccola superficie, sono sottili, ricche di capillari e mitocondri e caratterizzate da un elevato metabolismo ossidativo e dalla contrazione lenta (circa 100 ms). Si distinguono per una notevole resistenza alla fatica e un basso tono. Se un atleta possiede per propria costituzione una elevata percentuale di queste fibre, può essere favorito negli sport di durata.
- Fibre rapide (tipo II, bianche), ovvero “fast twitch fibers”, vale a dire fibre grandi con ampie superfici, spesse, povere di capillari e mitocondri, operanti più su base glicolitica che ossidativa; si contraggono rapidamente (circa 40-90 ms) ed altrettanto rapidamente si affaticano, sono inoltre caratterizzate da un tono elevato.
Sono a loro volta suddivise in:
a) Fibre di tipo IIA (fast twich oxidative fibers), esse possiedono un alto potenziale ossidativo ed un medio potere glicolitico. Utilizzano quindi un misto fra metabolismo aerobico ed anaerobico, sono relativamente resistenti alla fatica.
b) Fibre di tipo IIB (fast twitch glycolitic fibers), sono le fibre rapide per eccellenza, infatti esse possiedono unicamente un metabolismo di tipo anaerobico.
c) Fibre di tipo IIC, sono considerate intermedie, poco differenziate e sono modificabili, in un certo numero, con l’allenamento.
Ricapitolando…
Fibre di tipo I: lente, rosse, danno resistenza, dette anche fibre SO
Fibre di tipo II: bianche, rapide, danno forza/potenza/velocità
Fibre di tipo IIA: dette anche fibre FOG
Fibre di tipo IIB: dette fibre FG
Fibre di tipo IIC: fibre bianche, intermedie
Indipendentemente dalla velocità di contrazione, l’apparato contrattile dei vari tipi di fibre riesce a trasformare l’energia chimica in energia meccanica con un diverso grado di efficacia. In altre parole, il rapporto tra l’energia chimica utilizzata da un muscolo e l’energia meccanica da esso prodotta non non è sempre lo stesso.
In generale, nelle fibre di tipo I (a contrazione lenta) abbiamo una maggiore efficenza rispetto a quello di tipo II (a contrazione rapida), le quali hanno un elevato grado di eccitabilità. Le prime consumano meno energia, quindi si affaticano più lentamente delle altre. Inoltre, è importante ricordare che, a volte, fibre bianche e rosse si trovino distribuite nei diversi capi di uno stesso muscolo: nel tricipite, ad esempio, i capi brevi possono contenere in prevalenza fibre rosse, mentre le fibre bianche possono essere concentrate nel capo lungo.
La distribuzione delle fibre è perlopiù un fattore genetico. Tuttavia, delle biopsie, sembrano aver confermato che, nei soggetti non allenati, la percentuale di fibre bianche e rosse sia abbastanza simile (distribuzione abbastanza uniforme), negli atleti invece, l’allenamento costante produce una vera e propria specializzazione delle fibre, a seconda della disciplina praticata.
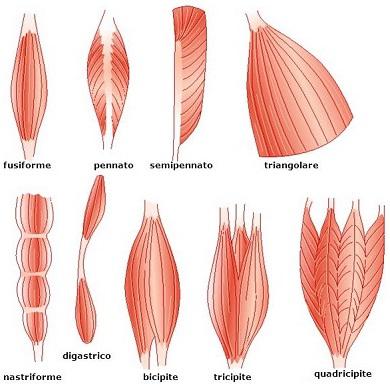
Forma
La forma dei muscoli scheletrici dipende dallo spazio in cui sono collocati, dalla disposizione interna delle fibre e dal tipo di lavoro a cui sono destinati. A seconda quindi della forma, i muscoli scheletrici sono suddivisibili nel seguente modo:
- fusiformi: le fibre sono parallele alla lunghezza;
- pennati: le fibre sono disposte obliquamente sui due lati del tendine;
- semipennati: le fibre sono disposte su un solo lato del tendine;
- pluripennati (o multipennati): sono quelli che hanno molti tendini di origine sui quali vanno a confluire e a tendersi le fibre muscolari;
- triangolari: sono appiattiti e con un piccolo tendine ad un’estremità;
- nastriformi: le fibre sono parallele alla lunghezza e interrotte da più inserzioni tendinee;
- digastrici: hanno un tratto tendineo intermedio tra i ventri muscolari.
A seguire, le illustrazioni delle varie tipologie di muscoli.

Contrazione muscolare (approfondimento)
La struttura miosinica e actinica, per effetto del processo di “scivolamento” dei filamenti, produce la contrazione muscolare. Come osservato nei primi anni 50 [1,2,3], l’accorciamento di questi due filamenti produce l’accorciamento del muscolo. La lunghezza dei filamenti spessi (miosina) non varia durante la contrazione, quella del sarcomero invece, si riduce (i filamenti spessi e sottili scivolano l’uno sull’altro). La forza di contrazione si sviluppa nel corso dello scorrimento di un tipo di filamento sull’altro.
La velocità di contrazione di una fibra dipende oltre che dalla velocità intrinseca di ogni sarcomero anche dal tipo predominante di miosina espressa (la molecola di miosina è composta da due molecole pesanti, 220 Kda).
Le varie tappe del processo di contrazione sono le seguenti:
- L’impulso nervoso arriva al muscolo attraverso la giunzione neuromuscolare sinaptica, eccitando le membrane che rivestono le miofibrille;
- in seguito a questo processo di eccitazione, si ha il trasferimento degli ioni calcio dall’esterno all’interno del sarcolemma;
- attraverso il sarcolemma, gli ioni calcio vanno a legarsi, da un lato, con i filamenti di actina e, dall’altro, con i filamenti di miosina staccando il ponte dal filamento di miosina. Va ricordato che l’actina e la miosina sono caricate negativamente lungo i loro filamenti e soltanto la miosina nell’estremità libera dei ponti presenta carica positiva;
- il calcio, neutralizzando due cariche negative, rompe l’equilibrio del muscolo a riposo dando la possibilità ai ponti di avvicinarsi alla base d’impianto perchè non più respinti;
- per effetto di questo avvicinamento, i filamenti di actina legati al ponte si uniscono a quelli della miosina;
- le miofibrille, a questo punto, sono soggette all’azione di un enzima detto ATP-asi (o adenosintrifosfatasi) che scinde l’ATP in ADP spezzando il collegamento che si era formato tra miosina ed actina;
- per l’azione di tale enzima, l’ADP viene nuovamente ricaricato in ATP; il ponte si ritrova ad avere una carica negativa all’estremità libera che viene nuovamente respinta sia dall’actina che dalla miosina.
Ipertrofia
Una delle caratteristiche più appariscenti dell’adattamento all’allenamento, specialmente per quanto riguarda quello con i sovraccarichi, è l’aumento del volume dei muscoli, fenomeno conosciuto come ipertrofia, definibile come l’incremento del materiale biologico che compone la struttura contrattile muscolare.
Esistono due tipi di ipertrofia:
- Ipertrofia sarcoplasmatica, caratterizzata da un incremento del volume sarcoplasmatico, dalla numerosità degli organuli cellulari contenuti al suo interno e dal rafforzamento delle strutture proteiche di sostegno che mantengono in asse le proteine contrattili e isolano l’ambiente intrafibrillare dall’ambiente esterno. Tale tipo di ipertrofia, stimolata da lavori non-massimali, ha discreti effetti positivi sulla produzione di forza muscolare;
- Ipertrofia miofibrillare, caratterizzata dall’incremento della densità delle proteine contrattili che compongono la testa della miosina e i siti attivi dell’actina. Tale incremento è controllato geneticamente dai mionuclei maturi della cellula (la cellula muscolare “polinucleata”, è la fusione specializzata di più cellule “mononucleate”). L’allenamento indirizzato allo sviluppo di forza massimale induce produzione di nuove proteine contrattili.
E in più: iperplasia (miofibrillare), caratterizzata dalla proliferazione di miocellule in serie o in parallelo attivate da cellule staminali in stato di quiescenza da fattori di crescita stimolati da lavori ad alta intensità.
L’ipertrofia muscolare inizia con l’aumento della matrice sarcoplasmatica, di una quantità proporzionale al numero di mionuclei presenti lungo la cellula muscolare. Ogni mionucleo è in grado di gestire la sintesi proteica stimolata 24-48h dopo l’allenamento in un’area di circa 2000 µm2. Ogniqualvolta la sezione trasversale muscolare gestita dal mionucleo aumenta, nuovi mionuclei vengono proliferati per garantire per ogni mionucleo una sezione muscolare ottimale.
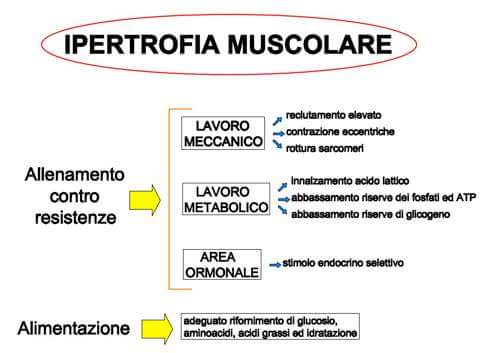
E’ importante tenere bene a mente alcuni punti riguardanti l’ipertrofia:
- essa è l’esito della supercompensazione del catabolismo proteico
- un carico molto alto comporta un’elevata intensità di catabolismo proteico ma limita il volume di lavoro e quindi le proteine catabolizzate in totale
- un carico molto basso permette un volume di lavoro notevole ma l’intensità dello stimolo non è tale da determinare un livello di catabolismo proteico apprezzabile
- le fibre rosse (lente, tipo I) sono poco ipertrofizzabili rispetto a quelle bianche (veloci, tipo II) ed ii risultati ottenuti, senza un frequente stimolo allenante, sono poco duraturi.
In una meta-analisi presente nella letteratura scientifica [4] si sono raggruppati in modo trasversale gli effetti iperplastici di differenti tipologie di protocolli di allenamento in diverse specie animali. I confronti hanno permesso di osservare ciò che per la maggior parte dei casi lo sviluppo muscolare è dovuto all’iperplasia e che il fenomeno dell’iperplasia muscolare longitudinale è maggiormente correlato ai protocolli di allenamento in cui si sviluppavano carichi eccentrici. Tuttavia, ciò che si verifica su alcuni animali non necessariamente è valido anche sull’essere umano. Infatti, non è attualmente chiaro se l’allenamento con i sovraccarichi stimoli o meno l’iperplasia muscolare. Senza l’utilizzo di steroidi anabolizzanti, quindi su soggetti natural, si crede che questo fenomeno sia poco rilevante [5].
Se accettate un consiglio spensierato, è meglio allenarsi per bene, senza troppe paranoie su iperplasia o non iperplasia.
Grazie per l’attenzione.

Bibliografia
Schoenfeld B. J. – The mechanisms of muscle hypertrophy and their application to resistance training (2010)
Urso A. – Le basi dell’allenamento sportivo (Calzetti Mariucci; 2a ediz., 2014)
Willmore H. J., Costill L. D. – Fisiologia dell’esercizio fisico e dello sport (Calzetti Mariucci, 2005)
D’Isep R., Gollin M. – Fitness e muscolazione (E.S.L., 2002)
1 Huxley A. F. & Niedergerke R. – Nature (1954)
2 H. E. Huxley and Hanson J. – ibid (1957)
3 H. E. Huxley – Science (1969)
4 Stone M. H. et al. – Principles and practice of Resistance Training (2007)
5 McDonald L. – Hyperplasia vs. Hypertrophy in Skeletal Muscle (2016)

Lascia un commento